This is the final post in my current series on memes, cultural evolution, and the thought of Daniel Dennett. You can download a PDF of the whole series HERE. The abstract and introduction are below.
* * * * *
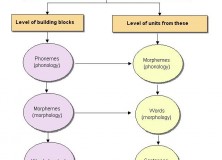
Abstract: Philosopher Dan Dennett’s conception of the active meme, moving about from brain to brain, is physically impossible and conceptually empty. It amounts to cultural preformationism. As the cultural analogue to genes, memes are best characterized as the culturally active properties of things, events, and processes in the external world. Memes are physically embodied in a substrate. The cultural analogue to the phenotype can be called an ideotype; ideotypes are mental entities existing in the minds of individual humans. Memes serve as targets for designing and fabricating artifacts, as couplers to synchronize and coordinate human interaction, and as designators (Saussaurian signifiers). Cultural change is driven by the movement of memes between populations with significantly different cultural practices understood through different populations of ideotypes.
* * * * *
Introduction: Taming the Wild Meme
These notes contain my most recent thinking on cultural evolution, an interest that goes back to my dissertation days in the 1970s at the State University of New York at Buffalo. My dissertation, Cognitive Science and Literary Theory (1978), included a chapter on narrative, “From Ape to Essence and the Evolution of Tales,” (subsequently published as “The Evolution of Narrative and the Self”). But that early work didn’t focus on the process of cultural evolution. Rather, it was about the unfolding of ever more sophisticated cultural forms–an interest I shared with my teacher, the late David G. Hays.
My current line of investigation is very much about process, the standard evolutionary process of random variation and selective retention as applied to cultural forms, rather than living forms. I began that work in the mid-1990s and took my cue from Hays, as I explain in the section below, “What’s a meme? Where I got my conception”. At the end of the decade I had drafted a book on music, Beethoven’s Anvil: Music in Mind and Culture (Basic 2001), in which I arrived at pretty much my current conception, but only with respect to music: music memes are the culturally active properties musical sound.
I didn’t generalize the argument to language until I prepared a series of posts conceived as background to a (rather long and detailed) post I wrote for the National Humanities Institute in 2010: Cultural Evolution: A Vehicle for Cooperative Interaction Between the Sciences and the Humanities (PDF HERE). But I didn’t actually advance this conception in that post. Rather, I tucked it into an extensive series of background posts that I posted at New Savanna prior to posting my main article. That’s where, using the emic/etic distinction, I first advanced the completely general idea that memes are observable properties of objects and things that are culturally active. I’ve collected that series of posts into a single downloadable PDF: The Evolution of Human Culture: Some Notes Prepared for the National Humanities Center, Version 2.
But I still had doubts about that position. Though the last three of those background posts were about language, I still had reservations. The problem was meaning: If that conception was correct, then word meanings could not possibly be memetic. Did I really want to argue that?
The upshot of this current series of notes is that, yes, I really want to argue it. And I have done at some length while using several articles by the philosopher Daniel Dennett as my foil. For the most part I focus on figuring out what kinds of entities play the role of memes, but toward the end, “Cultural Evolution, So What?”, I have a few remarks about long-term dynamics, that is, about cultural change. Continue reading “Cultural Evolution, Memes, and the Trouble with Dan Dennett”
About two years ago Wintz placed a comment on Replicated Typo’s About page in which he lists several papers that make good background reading for someone new to the study of linguistic and cultural evolution. I’ve just blitzed my way through one of them, Language is a Complex Adaptive System (PDF) by Beckner et al (2009)*, and have selected some excerpts for comment.
The point of this exercise is to contrast the way things look to a young scholar starting out now with the way they would have looked to a scholar starting out back in the ancient days of the 1960s, which is when both Dennett and I started out (though he’s a few years older than I am). The obvious difference is that, for all practical purposes, there was no evolutionary study of language at the time. Historical linguistics, yes; evolutionary, no. So what I’m really contrasting is the way language looks now in view of evolutionary considerations and the way it looked back then in the wake of the so-called Chomsky revolution—which, of course, is still reverberating.**
Dennett’s thinking about cultural evolution, and memetics, is still grounded in the way things looked back then, the era of top-down, rule-based, hand-coded AI systems, also known as Good Old-Fashioned AI (GOFAI). In a recent interview he’s admitted that something was fundamentally wrong with that approach. He’s realized that individual neurons really cannot be treated as simple logical switches, but rather must be treated as quasi-autonomous sources of agency with some internal complexity. Alas, he doesn’t quite know what to do about it (I discuss this interview in Watch Out, Dan Dennett, Your Mind’s Changing Up on You!). I’m certainly not going to claim that I’ve got it figured out, I don’t. Nor am I aware of anyone that makes such a claim. But a number of us have been operating from assumptions quite different from those embodied in GOFAI and Language is a Complex Adaptive System gives a good précis of how the world looks from those different assumptions. Continue reading “Dennett Upside Down Cake: Thinking About Language Evolution in the 21st Century”
I’ve already written one post in which I express skepticism about information-talk: Culture Memes Information WTF! I fear, alas, that it’s time for another. It’s not that I’m not aware of the concept of information, or that I haven’t made use of the concept, both the Shannon-Weaver technical concept and the more informal concept. But I think information talk is tricky.
As I said in that earlier post, a signal can be said to contain information only with respect to a system that can read and write that information. It’s one thing to talk about information when you have some technical understanding of the read-write mechanism, which seems to be the case in biology. But where such understanding is weak and vaporous, as it is in the case of human culture (the brain is more of a mystery than not) information talk is dangerous. Discussions of cultural evolution over the past quarter of a century or so have, for the most part, been blissfully uninformed by the cognitive and neurosciences and so, I fear, information talk has been a device for avoiding problems rather than solving them.
I get the sense that such talk is based on an underlying notion of pushing bits through a tube—cognitive linguists talk about the conduit metaphor. The nature of the tube doesn’t matter; its size, shape, substance, are all irrelevant. All that matters is the bits.
Language, for example, doesn’t work like that. All that goes through the tube is a physical signal—a sound wave, a visual signal (written or inscribed marks in the case of writing, gestures in the case of sign language), or a tactile signal (e.g. Braille). The meaning doesn’t go through the tube. The speaker’s intended meaning stays inside the speaker’s head. The listener constructs his or her own meaning according to his or her own perceptual and conceptual resources. In many cases these two meanings are congruent, especially in routine and relatively simple matters. There are, however, many cases where the two meanings ARE NOT congruent. In those cases congruence may be achieved through back-and-forth conversational negotiation. But not always, negotiations may fail. Continue reading “Information WTF 2: The Candy Itself”
On the face of it, Dennett and I have very different views about cultural evolution. To be sure, we both believe that Dawkins’s initial insight is valid: that culture is an evolutionary regime unto itself in which the benefits of cultural success accrue to cultural entities, not human individuals or populations. Where Dennett talks only of memes, I make an explicit distinction between memes and a cultural correlate of the phenotype (for which I have yet to adopt a term of art).
While Dennett allows memes to exist both in the external world and in the mind, most of his discussion is about memes in the mind moving from one mind to another. Indeed, I’d be curious to know what Dennett thinks exists in the mind apart from memes; of what, for example, does the neonate’s mind consist of? By contrast, I insist that memes exist in the external world, as observable (and memorable) properties of objects, events, and processes. The cultural correlates of the biological phenotype emerge as mental processes in brains as those brains engage with memes.
We thus have rather, if not utterly, different views about cultural evolution. As I have been thinking these things through, however, I have begun to suspect that our difference is more in how we assign roles in the process of cultural evolution to the mechanisms of human thought and action than in our conception of those mechanisms (though we no doubt have our differences there as well). And that’s the line I wish to investigate in this post. I will concentrate that investigation on a single essay:
Here’s a link to another conference that might be of interest:
The 3rd Linguistic Conference for Doctoral Students will take place at Heidelberg University, Germany from 05.-06. April 2013. The overarching topic of the conference will be: “Interdisciplinary Perspectives on Language, Discourse, and Culture.” The deadline for submissions is 15 February.
I’ve included the Call for Papers below (The Call for Papers can also be downloaded here):
For those of you who might be interested: Language and Cognition has a special issue on the nature and emergence of duality of patterning (paywall access, sorry!). As one of Hockett’s (1960) design features, duality of patterning is the property of human language that enables parts of language to be recombined in a systematic way to create new forms. In the introductory paper, de Boer, Sandler & Kirby (2012) identify two distinct levels where we see duality of patterning: combinatorial (meaningless sounds can be combined into meaningful morphemes and words) and compositional (morphemes and words can be combined to create new constructions with different meanings).For Hockett, not only is duality of patterning a design feature of language (in that all human languages have it), but also it is a unique characteristic of human language.
These two assumptions have been challenged on several fronts. First of all, simple combinatorial structure is found in systems of primate vocalisations, albeit restricted to a relatively limited set of signals. Meanwhile, in the Al-Sayyid Bedouin Sign Language (ABSL), the community does not have a conventionalised level of meaningless elements (although it does have compositional structure at the levels of morphology and syntax). These two examples offer important insights into the duality of patterning debate:
We see then from the case of ABSL that the need to express a large set of signals does not necessarily lead to combinatorial structure, while conversely from the animal systems, it appears that combinatorial structure does not necessarily need a very large set of signals to emerge. As combinatorial structure is the main defining characteristic of duality of patterning, it appears that both the status of duality of patterning as a design feature of language and the evolutionary pathways leading to it need to be rethought. (de Boer, Sandler & Kirby, 2012: 252).
The rest of the special issue is divided up between theoretical and experimental/modelling contributions. The abstracts and links to the papers (again, paywall, sorry!) are posted below. In summary, the general picture emerging from these papers is that duality of patterning is not a clearcut design feature of language, and nor is it necessarily a unique property of our capacity for language. Furthermore, we should also show a greater appreciation of the role that cultural evolution plays:
An apparent point of consensus from the papers in this special issue is that we should not see duality of patterning as a feature hard-wired into an innate language faculty, but rather as arising from multiple pressures operating on language as it emerges and changes in socially interacting populations. When we talk about the evolution of this design of language, then, we are referring more to cultural rather than biological evolution […] It appears that duality of patterning is a rather general state towards which sufficiently complex systems of signals evolve for different reasons: distinctiveness, learnability and a tendency to keep meaningful distinctions, while at the same time trying to make one’s utterances sound similar to those of others in the population. Thus, multiple cognitive processes seem to lead to duality of patterning and therefore, there are probably multiple evolutionary pathways that lead to duality of patterning as well. (de Boer, Sandler & Kirby, 2012: 257).
Stefan L. Frank, Rens Bod and Morten H. Christiansen
Abstract: It is generally assumed that hierarchical phrase structure plays a central role in human language. However, considerations of simplicity and evolutionary continuity suggest that hierarchical structure should not be invoked too hastily. Indeed, recent neurophysiological, behavioural and computational studies show that sequential sentence structure has considerable explanatory power and that hierarchical processing is often not involved. In this paper, we review evidence from the recent literature supporting the hypothesis that sequential structure may be fundamental to the comprehension, production and acquisition of human language. Moreover, we provide a preliminary sketch outlining a non-hierarchical model of language use and discuss its implications and testable predictions. If linguistic phenomena can be explained by sequential rather than hierarchical structure, this will have considerable impact in a wide range of fields, such as linguistics, ethology, cognitive neuroscience, psychology and computer science.
Published online before print September 12, 2012, doi: 10.1098/rspb.2012.1741 Proceedings of the Royal Society B
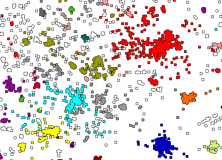
Gerhard Jäger uses lexostatistics to demonstrate that language similarities can be computed without using tree-based representations (for why this might be important, see Kevin’s post on reconstructing linguistic phylogenies). On the way, he automatically derives a tree of phoneme similarity directly from word lists. The result is an alternative and intuitive look at how languages are related (see graphs below). I review the method, then suggest one way it could get away from prior categorisations entirely.
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
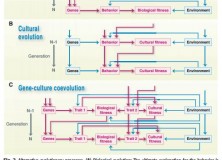
Figure 2 from Laland et al. (2011)
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.
Do you have a better memory than a chimp? Tetsuro Matsuzawa demonstrated the amazing working memory abilities of Chimpanzees, but maybe humans can be just as good, with enough practice. Justin Quillinan and I present the Quick-Hold Improvement Challenge (or QHImp Qhallenge). Play our game and find out if you can beat a chimpanzee.
You can see the results update live here. Results so far are tantalizing: Replicated Typo’s very own James Winters has already reached the Ayumu benqhmark (9 numbers viewed for only 209 milliseconds)!
Tetsuro Matsuzawa presented his work on chimpanzees in a plenary talk at Evolang. Matsuzawa covered several very interesting experiments and findings, including an experiment into the working memory of chimpanzees. Ayumu is a chimpanzee who was trained to recognise Arabic numerals on a touch-screen and press them in sequence. The most impressive aspect was that Ayumu could complete the task even when the numbers were only displayed for 210 milliseconds before being masked (the ‘eidetic memory task’ or ‘limited-hold’ memory task, Inoue & Matsuzawa, 2007):