About two years ago Wintz placed a comment on Replicated Typo’s About page in which he lists several papers that make good background reading for someone new to the study of linguistic and cultural evolution. I’ve just blitzed my way through one of them, Language is a Complex Adaptive System (PDF) by Beckner et al (2009)*, and have selected some excerpts for comment.
The point of this exercise is to contrast the way things look to a young scholar starting out now with the way they would have looked to a scholar starting out back in the ancient days of the 1960s, which is when both Dennett and I started out (though he’s a few years older than I am). The obvious difference is that, for all practical purposes, there was no evolutionary study of language at the time. Historical linguistics, yes; evolutionary, no. So what I’m really contrasting is the way language looks now in view of evolutionary considerations and the way it looked back then in the wake of the so-called Chomsky revolution—which, of course, is still reverberating.**
Dennett’s thinking about cultural evolution, and memetics, is still grounded in the way things looked back then, the era of top-down, rule-based, hand-coded AI systems, also known as Good Old-Fashioned AI (GOFAI). In a recent interview he’s admitted that something was fundamentally wrong with that approach. He’s realized that individual neurons really cannot be treated as simple logical switches, but rather must be treated as quasi-autonomous sources of agency with some internal complexity. Alas, he doesn’t quite know what to do about it (I discuss this interview in Watch Out, Dan Dennett, Your Mind’s Changing Up on You!). I’m certainly not going to claim that I’ve got it figured out, I don’t. Nor am I aware of anyone that makes such a claim. But a number of us have been operating from assumptions quite different from those embodied in GOFAI and Language is a Complex Adaptive System gives a good précis of how the world looks from those different assumptions. Continue reading “Dennett Upside Down Cake: Thinking About Language Evolution in the 21st Century”
For those doing or training to do academic research on linguistic and/or cultural evolution: Do memes matter to you?
I’ve got the impression that the issue that I’ve been chewing on recently, the appropriate account of memes of, if you prefer, the cultural analog of the biological gene, is mostly a theoretical one and has, so far, little bearing on empirical issues. However, I’ve also got the impression that most of the work on cultural evolution in the past decade or so has been empirical, either analysis of real-world data of one kind or another, or running simulations, and that the appropriate definition of meme doesn’t matter. You count what you can count. What matters is the quality of the raw data and the quality of the analysis.
I’ve been reading around in Dan Dennett’s papers and found this one, The Cultural Evolution of Words and Other Thinking Tools (Cold Spring Harbor Symp Quant Biol, Vol. LXXIV, August, 2009). To be sure, I disagree with his use of the meme concept. To be sure, his use is pretty standard and Dennett, in the standard way, claims more for it than can be justified by the current state of our knowledge and theorizing, but this paper is excellent despite that problem.
As the title indicates, Dennett focuses his attention on words and does so in a way that usefully brings their mystery, if you will, though mystery is rather low on Dennett’s intellectual agenda.
What then are words? Do they even exist? This might seem to be a fatuous philosophical question, composed as it is of the very items it asks about, but it is, in fact, exactly as serious and contentious as the claim that genes do or do not really exist. Yes, of course, there are sequences of nucleotides on DNA molecules, but does the concept of a gene actually succeed (in any of its rival formulations) in finding a perspicuous rendering of the important patterns amidst all that molecular complexity? If so, there are genes; if not, then genes will in due course get thrown on the trash heap of science along with phlogiston and the ether, no matter how robust and obviously existing they seem to us today.
For what it’s worth, I have it on good authority that there are languages which lack a word corresponding to our concept of word, though they generally have a word roughly corresponding to our concept of utterance (you can find this observation in, e.g., Alfred Lord, The Singer of Tales). That doesn’t bear directly on the point Dennett is making in those words as lacking a word for this is that really existing phenomenon is common enough, but it does indicate that words do have a rather diffuse or abstract character that makes it difficult to understand what they are and how they operate.
A bit later Dennett continues:
A promise or a libel or a poem is identified by the words that compose it, not by the trails of ink or bursts of sound that secure the occurrence of those words. Words themselves have physical “tokens” (composed of uttered or heard phonemes, seen in trails of ink or glass tubes of excited neon or grooves carved in marble), and so do genes, but these tokens are a relatively superficial part or aspect of these remarkable information structures, capable of being replicated, combined into elaborate semantic complexes known as sentences, and capable in turn of provoking cognitive, emotional, and behavioral responses of tremendous power and subtly.
I particularly like his phrase in that first sentence, “the trails of ink or bursts of sound that secure the occurrence of those words.” That secure the occurence, that’s nice. “Anchor” might also work, that anchor the occurence of those words in an utterance or a written text, as though the ink or sound were a tether holding the airy nothings of meaning and syntax to the ground. Continue reading “Dan Dennett on Words in Cultural Evolution”
UPDATE: This paper is now a Trends in Cognitive Sciences Free Featured Article and is available for free here
Noam Chomsky, who infamously stated that the field of language evolution research is “a burgeoning literature, most of which in my view is total nonsense” (see, e.g. here), has a new paper on the topic in press (together with linguist Robert Berwick and neuroscientists Angela Friederici and Johan Bolhuis) called Evolution, brain, and the nature of language (here, unfortunately it’s behind a paywall).
Here’s the abstract:
Language serves as a cornerstone for human cognition, yet much about its evolution remains puzzling. Recent research on this question parallels Darwin’s attempt to explain both the unity of all species and their diversity. What has emerged from this research is that the unified nature of human language arises from a shared, species-specific computational ability. This ability has identifiable correlates in the brain and has remained fixed since the origin of language approximately 100 thousand years ago. Although songbirds share with humans a vocal imitation learning ability, with a similar underlying neural organization, language is uniquely human.
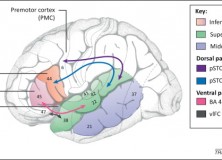
Also interesting is their figure on the Desing of the language system:
“The basic design of language. There are three components: syntactic rules and representations, which, together with lexical items, constitute the basis of the language system, and two interfaces through which mental expressions are connected to the external world (external sensory-motor interface) and to the internal mental world (internal conceptual-intentional interface).”
This still looks very much like the model advocated in for example, the influential and controversial Hauser/Chomsky/Fitch 2002 Science paper (see e.g. here) and from a brief look through the review. The paper also reiterates the view that language is primary an instrument aiding internal thought, and its use for communication is a later by-product (a view that has been thouroughly criticized, by for example Steven Pinker and Ray Jackendoff, e.g. here):
“communication, an element of externalization, is an ancillary aspect of language, not its key function, as maintained by what is perhaps a majority of scholars (cf. [Jim Hurford, Michael Tomasello], among many others). Rather, language serves primarily as an internal ‘instrument of thought’”
This post continues my summary of Jim Hurford’s discussion of two contrasting extreme positions on language evolution in his plenary talk at the Poznan Linguistic Meeting. Here’s the summary of these two positions from my last post:
Position A:
(1) There was a single biological mutation which (2) created a new unique cognitive domain, which then (3) immediately enabled the “unlimited command of complex structures via the computational operation of merge. (4) This domain is used primarily for advanced private thought and only derivatively for public communication. (5) It was not promoted by natural selection.
Position B:
(1) There were many cumulative mutations which (2) allowed the expanding interactions of pre-existing cognitive domains creating a new domain, which however is not characterized by principles unique to language. This then (3) gradually enabled the command of successively more complex structures. Also, on this view, this capacity was used primarily for public communication, and only derivatively for advanced private thought and was (5) promoted by natural selection.
Hurford criticized the position that the biological changes enabling languages primarily evolved for private thought, because this would imply that the first species in the Homo lineage that developed the capacity for unlimited combinatorial private thought (i.e. “merge”) were non-social and isolated clever hominids. This, as Hurford rightly points out, is quite unrealistic given everything we know about human evolution regarding, for example, competition, group size, neocortex side and tactical deception. There is in fact very strong evidence that what characterizes humans the most is the exact opposite as would be predicted by the “Merge developed in the service of enhancing private thought” position: We have the largest group size of any primate, the largest neocortex (which has been linked to the affordances of navigating a complex social world) and have the most pronounced capacity for tactical deception.
In his Talk, Hurford asked “What is wrong, and what is right, about current theories of language, in the light of evolution?” (you can find the abstract here).
Hurford presented two extreme positions on the evolution of language (which nevertheless are advocated by quite a number of evolutionary linguists) and then discussed what kinds of evidence and lines of reasoning support or seem to go against these positions.
Extreme position A, which basically is the Chomskyan position of Generative Grammar, holds that:
(1) There was a single biological mutation which (2) created a new unique cognitive domain, which then (3) immediately enabled the unlimited command of complex structures via the computational operation of merge. Further, according to this extreme position, (4) this domain is used primarily for advanced private thought and only derivatively for public communication and lastly (5) it was not promoted by natural selection.
On the other end of the spectrum there is extreme position B, which holds that:
(1) there were many cumulative mutations which (2) allowed the expanding interactions of pre-existing cognitive domains creating a new domain, which however is not characterized by principles unique to language. This then (3) gradually enabled the command of successively more complex structures. Also, on this view, this capacity was used primarily for public communication, and only derivatively for advanced private thought and was (5) promoted by natural selection.
Hurford then went on to discuss which of these individual points were more likely to capture what actually happened in the evolution of language.
He first looked at the debate over the role of natural selection in the evolution of language. In Generative Grammar there is a biological neurological mechanism or computational apparatus, called Universal Grammar (UG) by Chomsky, which determines what languages human infants could possibly acquire. In former Generative Paradigms, like the Government & Binding Approach of the 1980s, UG was thought to be extremely complex. What was more, some of these factors and structures seemed extremely arbitrary. Thus, from this perspective, it seemed inconceivable that they could have been selected for by natural selection. This is illustrated quite nicely in a famous quote by David Lightfoot:
“Subjacency has many virtues, but I am not sure that it could have increased the chances of having fruitful sex (Lightfoot 1991: 69)”
Here’s my opinion in a nutshell: This is a great volume and I’ve really learned a lot from reading it. The authors have done a great job trying to be accessible to an interdisciplinary audience. It’s a great place to start if you’re interested in language evolution or want to get a quick overview of a specific topic in language evolution research. I would’ve liked it if the chapters had a “Further Reading” section, however (like Christiansen and Kirby’s 2003 volume). Some chapters felt a bit too short for me (Steven Mithen‘s chapter on “Musicality and Language” for example is only 3 pages long, Merlin Donald‘s chapter on “the Mimetic Origins of Language” is 4 pages long). I also feel that some topics, like language acquisition, could’ve been dealt with more extensively, but then again, if you compile a handbook, it’s impossible to make everybody happy. Other recent book-length overviews of language evolution (e.g. Fitch’s 2010 book and Hurford’s 2007 and 2012 tomes) are more detailled, but also more technical and not as comprehensive and don’t cover as many topics. To quote my review:
Overall, the Oxford Handbook of Language Evolution is a landmark publication in the field that will serve as a useful guide and reference work through the entanglements and pitfalls of the language evolution jungle for both experienced scholars and newcomers alike.
One last thing I’m particularly unhappy about is that the handbook doesn’t have an Acacia Tree on the cover – which seems like a missed opportunity (kidding).
I’ll try to write about some of my favourite chapters in more detail somewhere down the road/in a couple of weeks.
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
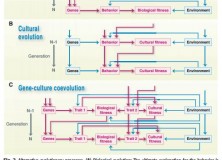
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
Figure 2 from Laland et al. (2011)
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.
The key to the treasure is the treasure.
– John Barth
In view of Sean’s post about Andrew Smith’s take on linguistic replicators I’ve decided to repost this rather longish note from New Savanna. I’d orignally posted it in the Summer of 2010 as part of a run-up to a post on cultural evolution for the National Humanities Center (USA); I’ve collected those notes into a downloadable PDF. Among other things the notes deal with William Croft’s notions (at least as they existed in 2000) and suggests that we’ll find language replicators on the emic side of the emic/etic distinction.
I’ve also appended some remarks I made to John Lawler in the subsequent discussion at New Savanna.
* * * * *
There’s been a fair amount of work done on language from an evolutionary point of view, which is not surprising, as historical linguistics has well-developed treatments of language lineages and taxonomy, the “stuff” of large-scale evolutionary investigation. While this work is directly relevant to a consideration of cultural evolution, however, I will not be reviewing or discussing it. For it doesn’t deal with the theoretical issues that most concern me in these posts, namely, a conceptualization of the genetic and phenotypic entities of culture. This literature is empirically oriented in a way that doesn’t depend on such matters.
The Arbitrariness of the Sign
In particular, I want to deal with the arbitrariness of the sign. Given my approach to memes, that arbitrariness would appear to eliminate the possibility that word meanings could have memetic status. For, as you may recall, I’ve defined memes to be perceptual properties – albeit sometimes very complex and abstract ones – of physical things and events. Memes can be defined over speech sounds, language gestures, or printed words, but not over the meanings of words. Note that by “meaning” I mean the mental or neural event that is the meaning of the word, what Saussure called the signified. I don’t mean the referent of the word, which, in many cases, but by no means all, would have perceptible physical properties. I mean the meaning, the mental event. In this conception, it would seem that that cannot be memetic.
That seems right to me. Language is different from music and drawing and painting and sculpture and dance, it plays a different role in human society and culture. On that basis one would expect it to come out fundamentally different on a memetic analysis.
This, of course, leaves us with a problem. If word meaning is not memetic, then how is it that we can use language to communicate, and very effectively over a wide range of cases? Not only language, of course, but everything that depends on language. Continue reading “In Search of the Wild Replicator”
Evolang is busy this year – 4 parallel sessions and over 50 posters. We’ll be posting a series of previews to help you decide what to go and see. If you’d like to post a preview of your work, get in touch and we’ll give you a guest slot.
Michael Pleyer Cognitive Construal, Mental Spaces, and the Evolution of Language and Cognition Poster Session 1, 17:20-19:20, “Hall” (2F), 14th March
Perspective-taking and -setting in language, cognition and interaction is crucial to the creation of meaning and to how people share knowledge and experiences. As I’ve already written about on this blog (e.g. here, here, here), it probably also played an important part in the story of how human language and cognition came to be. In my poster presentation I argue that a particular school of linguistic thought, Cognitive Linguistics (e.g. Croft & Cruse 2004; Evans & Green 2006; Geeraerts & Cuyckens 2007; Ungerer & Schmid 2006), has quite a lot to say about the structure and cognitive foundations of perspective-taking and -setting in language.
Therefore an interdisciplinary dialogue between Cognitive Linguistics and research on the evolution of language might prove highly profitable. To illustrate this point, I offer an example of one potential candidate for such an interdisciplinary dialogue, so-called Blending Theory (e.g. Fauconnier & Turner 2002), which, I argue, can serve as a useful model for the kind of representational apparatus that needed to evolve in the human lineage to support linguistic interaction. In this post I will not say much about Blending Theory (go see my poster for that 😉 or browse here ), but I want to elaborate a bit on Cognitive Linguistics and why it is a promising school of thought for language evolution research, something which I also elaborate on in my proceedings paper.
So what is Cognitive Linguistics?
Evans & Green (2006: 50), define Cognitive Linguistics as
“the study of language in a way that is compatible with what is known about the human mind, treating language as reflecting and revealing the mind.”
Cognitive Linguistics sees language as tightly integrated with human cognition. What is more, a core assumption of Cognitive Linguistics is that principles inherent in language can be seen as instantiations of more general principles of human cognition. This means that language is seen as drawing on mechanisms and principles that are not language-specific but general to cognition, like conceptualisation, categorization, entrenchment, routinization, and so forth.
From the point of view of the speaker, the most important function of language is that it expresses conceptualizations, i.e. mental representations. From the point of view of the hearer, linguistic utterances then serve as prompts for the dynamic construction of a mental representation. Crucially, this process of constructing a mental representation is fundamentally tied to human cognition and our knowledge of the world around us. Continue reading “Evolang Previews: Cognitive Construal, Mental Spaces, and the Evolution of Language and Cognition”














{kind=link}