I want to look at two recent pieces by Daniel Dennett. One is a formal paper from 2009, The Cultural Evolution of Words and Other Thinking Tools (Cold Spring Harbor Symposia on Quantitative Biology, Volume LXXIV, pp. 1-7, 2009). The other is an informal interview from January of 2013, The Normal Well-Tempered Mind. What interests me is how Dennett thinks about computation in these two pieces.
In the first piece Dennett seems to be using the standard-issue computational model/metaphor that he’s been using for decades, as have others. This is the notion of a so-called von Neumann machine with a single processor and a multi-layer top-down software architecture. In the second and more recent piece Dennett begins by asserting that, no, that’s not how the brain works, I was wrong. At the very end I suggest that the idea of the homuncular meme may have served Dennett as a bridge from the older to the more recent conception.
Words, Applets, and the Digital Computer
As everyone knows, Richard Dawkins coined the term “meme” as the cultural analogue to the biological gene, or alternatively, a virus. Dennett has been one of the most enthusiastic academic proponents of this idea. In his 2009 Cold Spring Harbor piece Dennett concentrates his attention on words as memes, perhaps the most important class of memes. Midway through the paper tells he us that “Words are not just like software viruses; they are software viruses, a fact that emerges quite uncontroversially once we adjust our understanding of computation and software.”
Those first two phrases, before the comma, assert a strong identification between words and software viruses. They are the same (kind of) thing. Then Dennett backs off. They are the same, providing of course, that “we adjust our understanding of computation and software.” Just how much adjusting is Dennett going to ask us to do?
This is made easier for our imaginations by the recent development of Java, the software language that can “run on any platform” and hence has moved to something like fixation in the ecology of the Internet. The intelligent composer of Java applets (small programs that are downloaded and run on individual computers attached to the Internet) does not need to know the hardware or operating system (Mac, PC, Linux, . . .) of the host computer because each computer downloads a Java Virtual Machine (JVM), designed to translate automatically between Java and the hardware, whatever it is.
The “platform” on which words “run” is, of course, the human brain, about which Dennett says nothing beyond asserting that it is there (a bit later). If you have some problems about the resemblance between brains and digital computers, Dennett is not going to say anything that will help you. What he does say, however, is interesting.
Notice that he refers to “the intelligent composer of Java applets.” That is, the programmer who writes those applets. Dennett knows, and will assert later on, that words are not “composed” in that way. They just happen in the normal course of language use in a community. In that respect, words are quite different from Java applets. Words ARE NOT explicitly designed; Java applets ARE. Those Java applets seem to have replaced computer viruses in Dennett’s exposition, for he never again refers to them, though they (viruses) figured emphatically in the topic sentence of this paragraph.
The JVM is “transparent” (users seldom if ever encounter it or even suspect its existence), automatically revised as needed, and (relatively) safe; it will not permit rogue software variants to commandeer your computer.
Computer viruses, depending on their purpose, may also be “transparent” to users, but, unlike Java applets, they may also commandeer your computer. And that’s not nice. Earlier Dennett had said:
Our paradigmatic memes, words, would seem to be mutualists par excellence, because language is so obviously useful, but we can bear in mind the possibility that some words may, for one reason or another, flourish despite their deleterious effects on this utility.
Once again, cultural evolution, and the problem of memes: What are they? Where are they? What do they do? While the general case does interest me, culture is so various that it is impossible to think about it directly. One has to think about specific cases. As details are important, I want to choose a fairly specific case, that of jazz in mid-20th-Century America. I want you to imagine that you’re in a jazz club in, say, Philadelphia, in, say, mid-October of 1952. It’s 1:30 in the morning, and the tune is Charlie Parker’s “Dexterity.” The piano player counts it off–ah one, ah two, one two three four…
But we’re getting ahead of ourselves. We need a little conceptual equipment before considering the example. It’s the conceptual equipment that’s in question. Make no mistake, the concept of memes is conceptual equipment, and it’s confused and confusing.
Roles in Cultural Selection
Genes and phenotypes play certain roles in a more or less standard account of biological evolution. The phenotype interacts with the environment, where it either succeeds or fails at reproduction, depending on the “fit” between its traits and that environment. Where the phenotype is successful at reproduction, it is the genes which are said to carry heredity from one generation to the next.
In one very widespread account genes are said to be replicators. That is to say, replication is the role they play in evolutionary change. Here’s what Peter Godfrey-Smith has to say about that (The Replicator in Retrospect, Biology and Philosophy 15 (2000): 403-423.):
In The Selfish Gene (1976), Richard Dawkins had argued that individual genes must be seen as the units of selection in evolutionary processes within sexual populations. This is primarily because the other possible candidates, notably whole organisms and groups, do not “replicate.” Organisms and groups are ephemeral, like clouds in the sky or dust storms in the desert. Only a replicator, which can figure in selective processes over many generations, can be a unit of selection.
At the same time Dawkins coined the term “meme” to name entities filling the replicator role in cultural evolution. Later on he used the term “vehicle” to designate the entity that interacts with the environment. In biological evolution it is phenotypes that are the vehicles. In cultural evolution, well, that’s a matter of some dispute. And that more general dispute–what are the roles in cultural evolution and what kinds of things occupy them?–is what interests me.
However, I don’t particularly like the term “vehicle.” As Godfrey-Smith has noted, following others, it is a gene-centric term, characterizing what entities do from the so-called “gene’s eye” perspective. I’d prefer a more neutral perspective and so will use a term coined by Richard Hull, “interactor.” Here are definitions as Godfrey-Smith gives them:
Still thinking about Dan Dennett’s conception of memetics. He’s got an article in the Encyclopedia of Evolution (Oxford 2005), “New Replicators, The” that’s worth looking at.
Some bits. From the beginning:
…evolution will occur whenever and wherever three conditions are met: replication, variation (mutation), and differential fitness (competition).
In Darwin’s own terms, if there is “descent [i.e., replication] with modification [variation]” and “a severe struggle for life” [competition], better-equipped descendants will prosper at the expense of their competitors. We know that a single material substrate, DNA (with its surrounding systems of gene expression and development), secures the first two conditions for life on earth; the third condition is secured by the finitude of the planet as well as more directly by uncounted environmental challenges.
The first question, then, is whether or not these conditions are met by human culture. Dennett thinks they are and so do I.
From the end, however:
Do any of these candidates for Darwinian replicator actually fulfill the three requirements in ways that permit evolutionary theory to explain phenomena not already explicable by the methods and theories of the traditional social sciences? Or does this Darwinian perspective provide only a relatively trivial unification?
We do not yet know. But are the prospects for non-triviality good enough to warrant considerable investment of conceptual time and energy? And so
We should also remind ourselves that, just as population genetics is no substitute for ecology—which investigates the complex interactions between phenotypes and environments that ultimate yield the fitness differences presupposed by genetics—no one should anticipate that a new science of memetics would overturn or replace all the existing models and explanations of cultural phenomena developed by the social sciences. It might, however, recast them in significant ways and provoke new inquiries in much the way genetics has inspired a flood of investigations in ecology. Continue reading “More on Dennett on Memes”
There is a “Skeptics In The Pub” event in Glasgow on March 4th, where Dr Thom Scott-Phillips will be discussing the perceptions and misconceptions of evolutionary psychology, in light of the public backlash against it that seems to be increasing all the time. This kind of public engagement is very sorely needed if we are to combat the rampant misinformation that crops up in both academic and non-academic communities. Among the criticisms being addressed at the event are the claims that evolutionary psychology is sexist, racist, or otherwise politically problematic. This is an important discussion.
From what I see around the feminist blogosphere, evolutionary psychology has a bad rap. Some recent examples I’ve come across include comments such as: “This new junk science named “evolutionary psychology” is the last variant of the male supremacy bible, following Freud’s mythology” and “[the way this article approaches the problem] is a bad idea [because] It smacks of evo psyche”. Even more liberal feminist blogs such as The F Word UK toe a similar line: Josephine Tsui seems to be on a personal mission against Evolutionary Psychology, armed with such ludicrous arguments as “You cannot replicate Evolutionary Psychology therefore it fails the methodologies of science” which display both an immature line of thinking and a fundamental misunderstanding of the theoretical motivations and methodologies entailed. Needless to say I’ve never seen this criticism leveraged against Evolutionary Biology, despite it being applicable to both.
Evolutionary psychology has a sound theoretical basis; it has been well established that natural selection is a means by which complex life and complex behaviour occurs. This tends to worry political movements like feminism, which has its roots in social constructionism. Such worry is unfounded; there is certainly a role for social constructionism within an evolutionary account of human behaviour. Put broadly, our plastic brains depend on complex social learning and pedagogy, which is an established cornerstone of human success. This ability to respond to (and be shaped by) the cultural environment has itself been selected for in humans, and can account for all manner of behaviours from language to mating preferences. Keep reading for a demonstration of how evolutionary psychology can in fact lend itself very well to the goal of engineering of social change.
So, on one side of the sexually selective understanding coin is a worried feminist movement, who risk losing a good grasp of evolutionary psychology by dismissing it entirely. On the other side, are the misogynist (mis)interpretations that have inspired this trepidation in the first place. That evolutionary psychology is abused and misinterpreted by misogynists and racists (and let’s be real here, this has happened a lot) is the problem, and it’s a serious one with real political consequences. Just this year, Steve Moxon submitted evidence to parliament (and was subsequently invited to speak) against the development of measures to encourage women in the workplace. Evolutionary psychology formed the backbone of his case, and he is not alone. Only an informed public can approach these claims with adequate discernment, so it is important that we address how some claims are morally wrong and incorrect. But it is also as important to discuss why they do not represent anything inherent about evolutionary psychology as a discipline.
We can illustrate the first way that evolutionary psychology can be wrong by using the problem of eugenics. Eugenics is theoretically sound, in as much as we know that we can selectively breed to a criteria and expect a predictable result; we’ve been doing it with dogs for 10 thousand years. This is also morally wrong and should not be attempted in humans. Just because eugenics is morally reprehensible, however, doesn’t mean we say the principles of artificial (or natural) selection aren’t true. Nor should this be the case for evolutionary psychology as a field; that it has been misapplied/misinterpreted within our social context (or just says something that we don’t like) simply does not speak to how scientifically correct it is. Another way that the interpretation of these studies can be grossly wrong is the Naturalistic Fallacy; the idea that if something is natural, it is inherently good or should be normative. This is obviously untrue; my human body is adapted to long-distance running, but I reject outright the idea that this is something I ought to engage in.
While citing the naturalistic fallacy is a good answer to most any claim about innate human proclivities, I think it’s also necessary to refute specific claims on their own grounds where possible. The final way for evolutionary psychology to be wrong is simply that rationalisation isn’t science, and instances where it is being passed off as such can be exposed for what they are. To illustrate, we generally do not dismiss the entirety of modern medicine as false because of the historical mistreatment of pregnant women in childbirth by doctors. Here, we can see that those occurrences are indeed morally wrong. However, it’s also the case that those instances are bad medicine by medicine’s own standards. Similarly, instances of bad science in evolutionary psychology, where latent misogyny and racism rears its head, can be refuted on their own grounds. This can and should be done without blithely dismissing the entire field.
It is a disaster that large factions of social justice movements are on the verge of outright anti-intellectualism when it comes to evolutionary psychology. Preserving ignorance about the field with out-of-hand dismissal neglects the potential for this tool to contribute to worthwhile political goals. We don’t have to stop at simply refuting the harmful instances of bad evolutionary study; there is also a positive agenda to be highlighted here. In the spirit of this, I’d like to share a preview of some work I’ve been hobbying with Justin Quillinan, inspired by a recent paper called “Asia’s Missing Women: A Problem in Applied Evolutionary Psychology?”. The paper aims to explore sex-preferential parental investment, which is a prolific problem in parts of Asia, the Middle East and North Africa, where the population’s sex-ratios are heavily male skewed as a result. It is already well documented that women suffer like this, so what can an evolutionary analysis can bring to the table? The problem, as presented in the paper, is this:
Asia’s missing women are, in economic terms, an aggregate outcome of millions of parenting decisions. The individual drivers behind those decisions emerge from interactions between our evolved parenting preferences and social and economic circumstances.
How do we untangle this seemingly nebulous problem? How do we determine why many self-perceived individuals act in such a similar way that the net result is the literal eradication of the female class? The approach of the paper seems to be largely influenced by economics, which is fairly central to a lot of evolutionary work. Game theory can be paraphrased as something like “given that the rules of the game are (x, y, z), which strategy should I employ to reap the most advantageous outcome?”. The most successful strategy is the one that is most likely to survive in the population, and hence it is the one we most expect to find. If we treat the problem of parents favouring boys over girls as a solution to the problem of parents’ circumstances, the question then becomes “what are the parameters that make this survival strategy worth employing such that it is so common?”
By comparing the commonalities in cultures that have this problem, Brooks identifies some ecomonic and social factors that may reward preferential parental investment. This is important: it means that campaigners for change don’t have to simply say the reason that girls are selectively aborted or neglected starts and ends with “girls are undervalued in these cultures”. Despite how true that is, it is also true of many cultures who do not have skewed sex ratios, and doesn’t really point to any concrete way of tackling the problem. If we can identify the driving factors that make parents behave this way with an evolutionary analysis, it means we can target specific structures with a specific end goal in mind.
In an interesting and wide-ranging investigation, the paper compares skewed parental investment occurence in non-human animals with the social and historical particulars that have led to this behaviour in disparate human populations. In doing so, Brooks proposes that male sex-biased populations are the systematic result of a population’s patrilineage, patrilocal kinship systems, and the dowry system. It was my hunch that the sex-biased population ratio could be reducible to patrilocality alone; that is, the system whereby women leave their blood relatives in order to live with (and care for) their husband’s family when they marry. Let us assume that the number of blood relatives in your family is a proxy measure of fitness. In a patrilocal social order, it is necessarily the case that having a son is more advantageous than having a daughter – precisely because daughters will always leave. Let us now also factor in the effects of infanticide and abortion; the option to neglect/kill your male/female offspring according to whether or not the most successful families you know had a boy or girl, will lead to the preferential elimination of females.
So we’ve implemented a model (source code available here) demonstrating exactly this:
We start with a population of agents separated into a number of families of a single breeding pair each single individuals. At each time-step, the following events occur: 1. Reproduction: fertile breeding pairs of agents have a new agent ‘child’ of random sex. A fertile pair is one that does not have an unmarried child that is younger than the age of maturity. 1.2. Abortion: Now, the pair can choose to either keep or to abort their new agent. To make this decision, they choose a random family, with larger families having a proportionally higher probability of being chosen. If the sex ratio of that family is a mismatch to the child they have just had, they will abort the child. Otherwise, they will keep it. (ETA: the abortion decision is based on the sex ratio of all the offspring of the chosen family). 2. Marriage: Every single, mature agent attempts to pair with a random, opposite sex partner from a different family, to form a breeding pair. In patrilocality, the female leaves her family and is appended to her husband-agent’s family to form a breeding pair. (In the matrilocality sanity check, the situation is vice versa and male agents join their wives’ families). 3. Death: Agents above a certain age are removed from the population. If a family no longer has any members, we generate a new breeding pair individual so that the population doesn’t die out.
The first null model was as above, with the omission of step 1.2. We later implemented one that is as above, but minus patrilocal marriage (ie, married agents simply form a new family pair) because this is a better comparison.
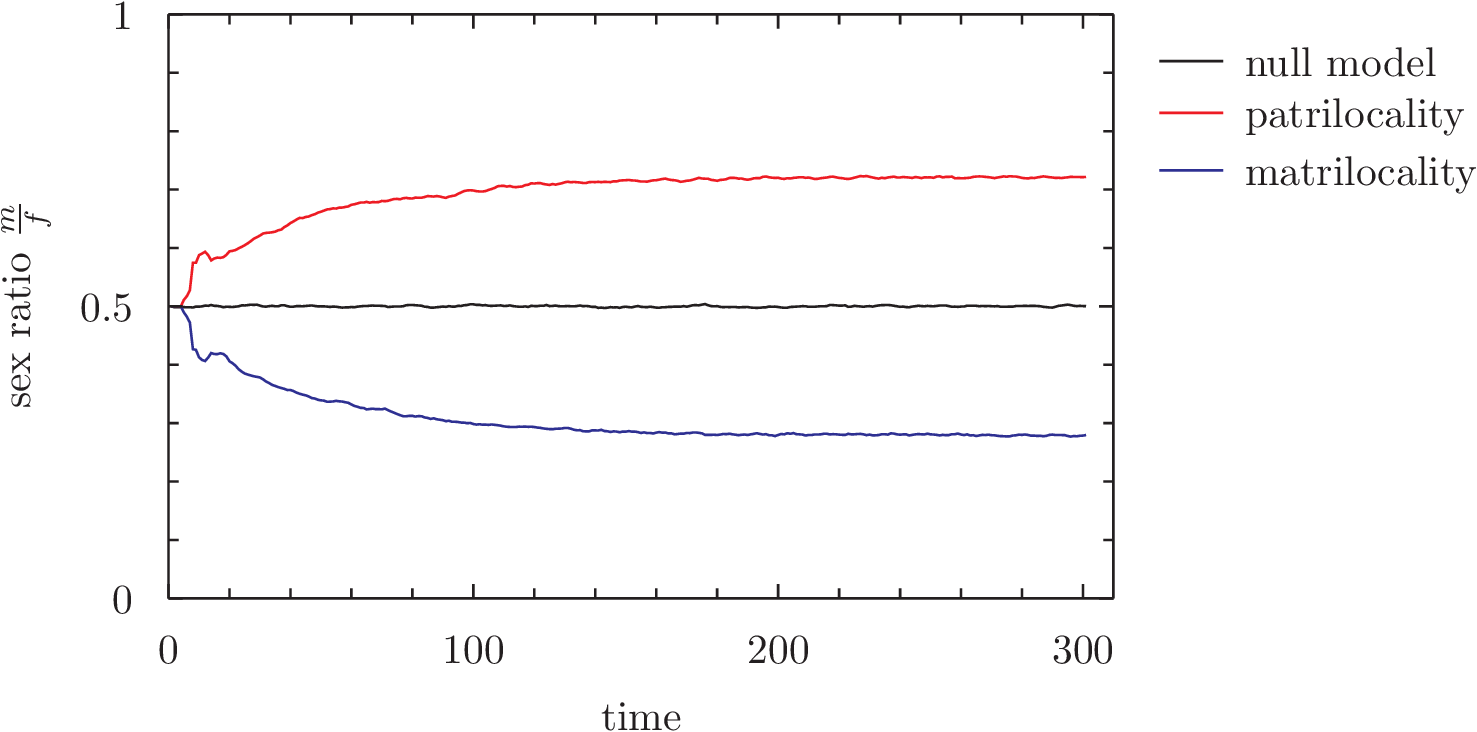
At each time-step, we measure the sex ratio of the population. This is what happens (wordpress is terrible, click for a clearer image):
Average of 500 runs, seeded from 50 family breeding pairs
The sex-ratio of the population is skewed in the direction of the sex that determines family locality – that is to say, patrilocality alone systematically results in the preferential abortion of female offspring, and a higher ratio of males in the population to females. This model will hopefully lend itself to some further work exploring the role of the dowry in maintaining the system by offsetting the costs of giving away offspring, preferential marrying, and how a shift toward “nuclear” family arrangements may have lifted the cost-benefit situations disadvantaging females (and thus making dowry systems redundant).
UPDATE (09/02/12): Here is the data using an amended null model:
Average of 1000 runs, seeded from 50 individuals
The ‘bump’ at around time-step 20 in the first graph noticed by Sean (see comments section) doesn’t appear here; this was an artefact of seeding with identical pairs that breed and die at the same time. Seeding with individuals has smoothed out that curve; staggering the ages of agents would likely smooth it out further. The extra noise in this model means that the skew is less pronounced than before; note the Y axis is zoomed in to ~0.3 – 0.7. The null model here is of the null hypothesis; abortion still happens, the only difference is that instead of a married agent appending to their spouse’s family, the married couple form a new family pair (ie. no matri/patrilocality). This means that any single given population’s sex ratio is susceptible to drift; early, small aberrations toward male or female will become magnified over time. This is, however, equally likely to happen for either male or female, and so the average of 1000 populations shown here is stable at 0.5.
Further implications:
An important additional observation in Brooks’ paper is an examination of the wider social consequences of this particular set of circumstances. The paper names elevated levels of “men competing furiously for wealth and status” as well as “risk-taking, violence, gambling, alcohol and drug abuse, kidnapping and trafficking of women, and the use and abuse of prostitutes” as consequences of surplus males in the population. The implication is that, by this model, these large societal problems can be addressed at least in part by balancing parental investment in children of both sexes, which would be remarkable.
At first blush, the idea that violence results from a surplus of men who don’t have a good enough chance at mating with women has some worrying and problematic implications. It is, nevertheless, intuitively true within a culture of male entitlement, which is something that feminists have long observed – that male violence is the result (and the maintenance) of a patriarchal social order. Since patrilineage, and patrilocality in family structure specifically, are identified as the preconditions for preferential parental investment in males, the eradication of this social order is a necessary step in redressing the sex-ratio balance. The end of patrilineal traditions and patrilocality are also a step toward dismantling a culture of male entitlement more broadly. As a direct consequence, then, this strategy dismantles the structures supporting male entitlement itself at the same time as addressing the skewed sex ratio, and does not simply consist of “giving the men more women to stop them fighting”.
It seems to me that a feminist account that names a culture of male entitlement as the cause for violent female oppression, and an evolutionary account that names structural entitlement systems as the cause for the mass devaluation/infanticide of female offspring are very much on the same page. This approach also very clearly illustrates the compatibility between evolutionary analysis and the socio-economic determinism that is fundamental to radical political thought, precisely by demonstrating how population-wide behaviour can directly result from external economic and social parameters, rather than some innately predisposed condition. We hope that this is at least one small demonstration of how evolutionary psychology and social justice can be rather natural allies.
Following on from the Baronchelli et al paper a couple of months ago, PLOS ONE has published “Evolution in a Changing Environment” by the same authors. The conclusions of the 2 papers both argue that if language is rapidly changing (and it is), then generalist, neutral genes, rather than specialist ones, are advantageous. This argues that language is likely more the result of general cognitive abilities as language change happens so rapidly. In contrast to the last paper though, this one focuses much less on (specifically) linguistic change, and features a super sexy stochastic interacting particle model (if you’re into that sort of thing).
Abstract:
We propose a simple model for genetic adaptation to a changing environment, describing a fitness landscape characterized by two maxima. One is associated with “specialist” individuals that are adapted to the environment; this maximum moves over time as the environment changes. The other maximum is static, and represents “generalist” individuals not affected by environmental changes. The rest of the landscape is occupied by “maladapted” individuals. Our analysis considers the evolution of these three subpopulations. Our main result is that, in presence of a sufficiently stable environmental feature, as in the case of an unchanging aspect of a physical habitat, specialists can dominate the population. By contrast, rapidly changing environmental features, such as language or cultural habits, are a moving target for the genes; here, generalists dominate, because the best evolutionary strategy is to adopt neutral alleles not specialized for any specific environment. The model we propose is based on simple assumptions about evolutionary dynamics and describes all possible scenarios in a non-trivial phase diagram. The approach provides a general framework to address such fundamental issues as the Baldwin effect, the biological basis for language, or the ecological consequences of a rapid climate change.
UPDATE: This paper is now a Trends in Cognitive Sciences Free Featured Article and is available for free here
Noam Chomsky, who infamously stated that the field of language evolution research is “a burgeoning literature, most of which in my view is total nonsense” (see, e.g. here), has a new paper on the topic in press (together with linguist Robert Berwick and neuroscientists Angela Friederici and Johan Bolhuis) called Evolution, brain, and the nature of language (here, unfortunately it’s behind a paywall).
Here’s the abstract:
Language serves as a cornerstone for human cognition, yet much about its evolution remains puzzling. Recent research on this question parallels Darwin’s attempt to explain both the unity of all species and their diversity. What has emerged from this research is that the unified nature of human language arises from a shared, species-specific computational ability. This ability has identifiable correlates in the brain and has remained fixed since the origin of language approximately 100 thousand years ago. Although songbirds share with humans a vocal imitation learning ability, with a similar underlying neural organization, language is uniquely human.
Also interesting is their figure on the Desing of the language system:
“The basic design of language. There are three components: syntactic rules and representations, which, together with lexical items, constitute the basis of the language system, and two interfaces through which mental expressions are connected to the external world (external sensory-motor interface) and to the internal mental world (internal conceptual-intentional interface).”
This still looks very much like the model advocated in for example, the influential and controversial Hauser/Chomsky/Fitch 2002 Science paper (see e.g. here) and from a brief look through the review. The paper also reiterates the view that language is primary an instrument aiding internal thought, and its use for communication is a later by-product (a view that has been thouroughly criticized, by for example Steven Pinker and Ray Jackendoff, e.g. here):
“communication, an element of externalization, is an ancillary aspect of language, not its key function, as maintained by what is perhaps a majority of scholars (cf. [Jim Hurford, Michael Tomasello], among many others). Rather, language serves primarily as an internal ‘instrument of thought’”
Sorry for the hyperbole in the title, but now I’ve got your attention – Researchers at the University of Edinburgh have found a gene which is implicated in human brain development which humans have, but chimpanzees don’t.
The study compared the human genome to 11 other species of mammals, including chimpanzees, gorillas, mice and rats, and found that miR-941 is unique to humans.
miR-941 is now being slated as a gene that contributed to how early humans developed tool use and language. This gene, in contrast to the likes of FoxP2, has a very specific function rather than being a gene that down regulates other functions. It is said to be the only gene discovered that has such a specific function while only being present in humans. It is active in two areas of the brain that control our linguistic abilities and also our decision making.
The authors estimate that it emerged between six and one million years ago and that it emerged fully functional out of non-coding genetic material (“junk DNA”) in a very short interval of evolutionary time.
I’m sure the studies where they implant it into mice will start soon. Watch this space.
Gerhard Jäger uses lexostatistics to demonstrate that language similarities can be computed without using tree-based representations (for why this might be important, see Kevin’s post on reconstructing linguistic phylogenies). On the way, he automatically derives a tree of phoneme similarity directly from word lists. The result is an alternative and intuitive look at how languages are related (see graphs below). I review the method, then suggest one way it could get away from prior categorisations entirely.
A forthcoming paper (grateful nod to ICCI) in PNAS from Olivier Mascaro and Gergely Csibra presents a series of experiments investigating the representation of social dominance relations in human infants, and it’s excellent news: we’re special.
Social dominance can be inferred in a couple of ways. Causal cues such as age, physical aggression and size can tell us about the dominance status of an individual quite intuitively, so we can make a sensible decision about whether or not we get into a scrap with them. Another way we can establish this is to look for direct realisations of dominance, such as who gets the banana if two hungry chimps both want it; chances are, little Pan Pipsqueak isn’t going to get a look in. In order to be useful, we also have to use this information to expect certain things from the individuals around us, so those representations have some property of stability across time that allows us to have those expectations. The question being explored in this paper is whether the representations we have are about the relationship between the two agents who want the banana, or the individual properties each of them has.
In a series of experiments using preferential looking time as a dependent measure, human infants (9 and 12 month olds) were exposed to videos of geometric figures exhibiting similar goal-directed behaviour. Then they would watch, say, a dominant triangle picking up the last figurative banana when the nondominant pentagon also wanted it. For expository purposes and posterity’s sake, I have constructed an artist’s impression of a dominant triangle and a subordinate pentagon in MSPaint (below, right):
A dominant triangle and subordinate pentagon (artist’s impression).
I’m not just showing off my extraordinary artistic talent here; the good thing about these agents is that there are none of the cues like size or aggression that can give rise to the assignment of individual dominance properties. The task also doesn’t indicate anything similar; it’s just about who gets the desired object when there’s only one left. In other words, the goal-directed actions of two agents are in opposition. After seeing a triangle beat a pentagon to an object of ‘banana’ status, 12 month olds looked for longer when they were then presented with an incongruent trial where the pentagon gained over the triangle. 9 month olds (understandably?) couldn’t care less. So, on the basis of this social interaction alone, the 12 month olds were able to notice when something unexpected happened.
To rule out the possibility that this was just the result of some simple heuristic such as “when triangle and pentagon are present, triangle gets the object” and make sure the infants really were assigning some dominance, another experiment (with 12 and 15 month olds) showed the same test video of the two agents collecting little objects. This time, however, the preceding video was of the triangle dominating a little walled-in space that the pentagon also wanted to inhabit. The 12 month olds had no idea what was up, but the 15 month olds generalised from the first “get out of my room” interaction to the “I get the last banana” interaction. So, 15 month olds can extract, just from watching a social interaction, the dominance status of agents and can generalise that information to novel situations. So if a 15 month old watches you lose your favourite seat in front of the TV, they’ll also expect you to miss out on the last slice of pizza, because you’re a loser.
What we still don’t know is whether they think your belly is inherently yellow, or if you’re just a pushover when interacting with a particular person. Is it the relationship between the triangle and pentagon that the babies are tracking, or do they just give each agent some sort of dominance score? This was addressed in experiment 4, where infants were presented with two interactions: one between A and B, where A wins, and then another between B and C, where B wins. If the babies are assigning an individual value to each agent, they should have some sort of linear, transitive representation of dominance like A > B > C. If they’re then presented with a novel interaction between A and C, they would have the expectation that A will beat C. So if they stare in surprise at a trial where C wins, we know it’s violated that kind of expectation, and that they’re representing this stuff linearly – I.E. each agent has a dominance value. In contrast, if the infant is tracking the relations between agents, they can’t really have an expectation of what will happen when A and C both want a banana, because they’ve never seen C before. The results find that the infants look preferentially when they get an incongruent trial using agent pairs they have seen before – as we’d expect from the previous experiment. When they’re presented with a new “I get the last banana” interaction between A and C, however, there’s nothing startling about it when C wins – which means their expectations are not based on something like A > B > C.
The only tiny little harrumph I have about this result is that all it does is falsify the linear representation account. Though I think their account is absolutely right, it’d be nice to see something more predictive come out of the relation-representation hypothesis that is a little more falsifiable. But this result is pretty huge, and stands in contrast with what we know about social cognition in other animals like baboons (Cheney et al, 1995; Bergman et al, 2003), lemurs (Maclean et al., 2008) and even pigeons (Lazareva & Wasserman, 2012), who seem to employ this sort of hierarchical, transitive inference when presented with novel interactions. It may also muddy the waters a little when we want to make the appealing claim that, since language surely emerged in order to enable communication as we navigated a social environment, hierarchical social cognition gives rise to the processing of languagey things like hierarchical syntax or our semantic representation (Hamilton, 2005), which can be characterised as hierarchical (e.g. hyperonym > hyponym). If we consider the nature of the human social environment, though, it should seem more intuitive that something more reliable than simple transitive inference is necessary in order to successfully navigate through our interactions. Due to our prolific production of (and reliance on) culture, humans have a much more diverse range of social currencies, which correspond to values for things like money, intelligence, blackmail information, who your friends are, ad infinitum. That means it’s pretty reasonable that our social cognition needs new strategies in order to get by; we have a little more to consider than just who’s big and angry enough to get all the bananas.
References
Bergman, T., Beehner, J., Cheney, D. & Seyfarth, R. (2003) “Hierarchical Classification by Rank and Kinship in Baboons” Science 14(302), 1234-1236.
Cheney, D., Seyfarth, R. & Silk, J. (1995) “The response of female baboons (Papio cynocephalus ursinus) to anomalous social interactions: evidence for causal reasoning?” Journal of Comparative Psychology 109(2), 134-141.
Lazareva, O. & Wasserman, E. (2012) “Transitive inference in pigeons: measuring the associative values of stimulus B and D” Behavioural Process 89(3), 244-255.
Maclean, E., Merritt, D. & Brannon, E.M. (2008) “Social complexity predicts transitive reasoning in prosimian primates” Animal Behaviour 76(2), 479-486.
Mascaro, O. & Csibra, G. (forthcoming) “Representation of stable dominance relations by human infants” Proceedings of the National Academy of Sciences
A somewhat contentious debate among the behavioural sciences is currently underway concerning Mayr’s division of causal explanations in evolutionary theory. Here I’m going to give you a brief rundown of two papers in particular, before I chip in my two-cents about how other insights from the theoretical literature can inform this debate. It seems the discussion is just getting started with respect to cultural evolution, so it’d be interesting to hear other peoples’ comments from either camp.
Over the years, evolutionary theorists have tried to make logical divisions between the kinds of things we can ask about, with a view to making it clear what exactly scientific studies can tell us. A dominant paradigm dividing two levels of causation for biological features we see in the world is Mayr’s distinction between ultimate and proximate causes. Ultimate causation explains the proliferation of a trait in a population in terms of the evolutionary forces acting on that trait. For example, peahens that prefer peacocks with larger tails (an honest signal of fitness following the handicap principle) will have stronger or more successful offspring, and so this preference proliferates along with larger peacock tails. Proximate causation uses immediate physiological and environmental factors to explain a particular peahen’s penchant for a large-tailed peacock in a mate choice trial, where the signal of the peacock’s large tail elevates the hormone levels in the peahen and copulatory behaviour ensues. Although the behaviour in both of these examples is the same, the levels of explanation are based on different sets of factors.
In Perspectives on Psychological Science last year, a paper by Scott-Phillips, Dickins and West voiced some concerns about these two levels of causation being conflated in the behavioural sciences. In particular, they addressed instances where proximate explanations of traits are being framed as ultimate ones. The paper points specifically to studies of the evolution of cooperation, transmitted culture and epigenetics to illustrate this. Regarding the evolution of cooperation, they point to an instance where ‘strong reciprocity’ (an individual’s propensity to reward cooperative norms and sanction violation of these norms) is purported to be an ultimate explanation of why humans cooperate, rather than a proximate mechanism that enables such cooperation.
Table 1 from Scott-Phillips et al. (2011), highlight added.
Among the examples was the feature of linguistic structure (see table 1 from paper above), where several studies pointed to the cultural transmission process as an ultimate explanation of linguistic structure. They suggest that cultural transmission constitutes a proximate process, because it gives the means by which linguistic structure is expressed – and this is how cultural transmission contributes to what the linguistic structure looks like. One analogy might be that the vibrating of my particular vocal cords is a proximate mechanism giving rise, in part, to how my voice sounds, rather than an ultimate explanation of why I vocalise. Since an ultimate account must suggest how a trait contributes to inclusive fitness in order to explain its prevalence in humans, they uncontroversially venture that the ultimate rationale for the ubiquity of linguistic structure is that it greater enables communication (and therefore increases inclusive fitness by enabling cooperative activity).
An opposing view was later published in Science by Laland, Sterelny, Odling-Smee et al., who suggest that the use of Mayr’s division of ultimate and proximate causation is not helpful to all evolutionary investigations, and even hampers progress. The grounds for rejecting Mayr’s paradigm seem to lie largely in what Laland et al. term “reciprocal causation”. That is, that “proximate mechanisms both shape and respond to selection, allowing developmental processes to feature in proximate and ultimate explanations”. After aligning proximate explanations with ontogeny and ultimate explanations with phylogeny, they suggest that what we may have called ultimate and proximate features are no longer sharply delineated, and that these reciprocal processes mean that the source of selection sometimes cannot be separated. They present an idea from the field of evolutionary-developmental biology that, if a developmental process makes some variant of a trait more likely to arise than others, then this proximate mechanism helps to construct an “evolutionary pathway”.
Figure 2 from Laland et al. (2011)
The paper also highlights developmental plasticity, and gene-environment interaction more broadly (see fig. 2 from paper, above), as a process where reciprocal causation offers an evolutionary explanation conceptually comparable to ultimate causation. Talking specifically on the topic of linguistic structure, they present the debate about whether specific design features of language are attributable to biological or cultural evolution. The paper points out that cultural evolution determines features of linguistic structure – for example, word order – and that the existing word order determines that of future speakers. Indeed, at the Edinburgh LEC we know that transmission by iterated inductive inference under general conditions can explain particular structures in languages. That cultural evolution determines the variation between languages, Laland et al. say, provides evidence that it is an evolutionary force comparable to natural selection (and, therefore, ultimate explanation).
What follows is a collection of my thoughts on the matter, which are (spoiler alert) largely in support of the Scott-Phillips et al. paper. I hope others more experienced in cultural evolution studies than I will contribute their perspective.
It seems to me that there are a few assumptions made in the Laland et al. paper that are not quite in line with how Mayr himself understood the paradigm. Perhaps much can be learned from this debate’s previous incarnation, when Richard C. Francis made similar arguments against the ultimate/proximate distinction in 1990. In his critique, he equated ultimate causation with phylogeny and proximate causation with ontogeny – an approach that was rebuked by Mayr in 1993, who made the point that “all physiological activities are proximately caused, but is a reflex an ontogenetic phenomenon?”. Mayr’s response is actually rather unhelpful in addressing the arguments fully, and this statement is particularly dense. But what he is getting at here is the idea that interaction with the environment that gives rise to adaptive behaviours (such as recoiling instantly from a hot stove) is itself subject to selection, and thus constitutes a proximate explanation of causation. Relatedly, he points out that most components of the phenotype are indeed the result of genetic contribution and interaction with the environment, which has been successfully explored in biology within the traditional theoretical paradigm.
A perhaps more nuanced account of how we can divide the possible explanations of biological phenomena is offered by Tinbergen in his “four questions”, where ultimate explanations are further subdivided into Function (concerning the adaptive solution to a survival problem favoured by natural selection) and Phylogeny, which is a historical account of when the trait arose in the species, and importantly includes processes other than natural selection that give rise to variation – such as mutation, drift and the constraints imposed by pre-existing traits (see blind spot example below). Proximate explanations are further split into Mechanism (immediate physiological/environmental factors causal in how the trait operates in the individual) and Ontogeny (the way in which this trait develops over the lifetime of the individual). As a simple example, here is the paradigm applied to a trait like mammalian vision that I lifted from Wikipedia: Ultimate Function: To find food and avoid danger. Phylogeny: The vertebrate eye initially developed with a blind spot, but the lack of adaptive intermediate forms prevented the loss of the blind spot. Proximate Causation: The lens of the eye focuses light on the retina Ontogeny: Neurons need the stimulation of light to wire the eye to the brain within a critical period (as those awful studies of blindfolded kittens illustrated).
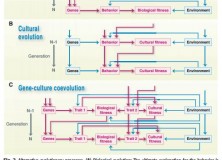
A schematic below, adapted from Tinbergen (1963) shows how these levels of causation may interact with one another, which appears to communicate something roughly comparable to the importance Laland et al. place on “reciprocal causation” in the formation of adaptive variants:
Adapted from Tinbergen (1963); Causal Relationships
Applied to the debate outlined above, it would seem that there is no apparent reason that a process of gene-environment interaction – including the cultural environment – can’t itself be subject to selection, or that developmental plasticity itself is not an adaptation in need of an ultimate explanation. It has long been the case that behaviour is no longer understood as either “nature” or “nurture”, but gene-environment interaction, with varying levels of heredity. The “reciprocal causation” suggested in Laland et al.’s paper, is (as they point out) very common in nature; feedback loops are uncontroversial proximate processes in biology. That a proximate process may give rise to a dominant variant of a trait in a population does not explain why it is adaptive, and this points to another problem with the proposing the abandonment of Mayr’s paradigm: a logical division of levels of explanation doesn’t seem to be the sort of thing that can be rendered outdated by empirical evidence. Indeed, claims about the particulars of traits and processes (and languages) themselves are a matter for empirical data – but the theoretical issue about the level of explanation that data is useful for does not itself seem to be subject to empirical findings.
The finding that a proximate process such as cultural transmission gives rise to a trait that is prolific in a population is itself exciting and surprising, and even shows us that the pressure for making language easier to learn gives us adaptive languages to learn; however, it could be argued that it is this process that is adaptive, and that the reason why humans so heavily rely on this process is an ultimate explanation.
One way of resolving these two perspectives may be to place cultural processes that give rise to variation at the level of what Tinbergen labels Phylogenetic (one subset of ultimate) explanation, as it concerns processes which produce some heightened frequency of traits over a language’s history. An explanation at the level of Phylogeny still must make recourse to natural selection at some point, since variants that result from mutation or drift are retained because of their adaptive value (or an adaptive trade-off). This approach may be a problem for the current understanding, which holds that the features resulting from cultural processes are themselves adaptive and therefore comparable to what Tinbergen labels Function.
The problem with this is that calling particular structures of language ‘adaptive’ obscures what it is about Language that is actually being selected for. To flesh out what I mean, I think it’s useful to consult Millikan’s (1993) distinction between Direct Proper Function and Derived Proper Function (… bear with me, it’ll be worth it, honest). The Direct Proper Function of a given trait T can be thought of as a “reproduction” of an item that has performed the exact same adaptive function F, and T exists because of these historical performances of F. Sperber and Origgi (2000) use the illustrative example of the heart, where the human heart has a bunch of properties (it pumps blood, makes a thumping noise, etc), but only its ability to pump blood is its Direct Proper Function. This is because even a heart that doesn’t work right or makes irregular thumping noises or whatever, still has the ability to pump blood. Hearts that pump blood have been “reproduced through organisms that, thanks in part to their owning a heart pumping blood, have had descendents similarly endowed with blood-pumping hearts”.
The Derived Proper Function, however, refers to a trait T that is the result of some device that, in some environment, has a Proper Function F. In that given environment, F is usually achieved by the production of something like T. If I unpack this idea and apply it to language, we can understand it as the acquisiton and production of a device that, in this environment, leads to, say, a particular SVO language, T. The Proper Function of adaptive communication is performed by T in this case, but could also be performed by any number of SOV, VSO, etc Ts in other cases. In other words, the Proper Function of this language is not the word order itself, but communication. The word order is the realisation of this device that is reproduced because of the performance of T in a particular environment, but does not necessarily lead to T in the next incarnation of that device (i.e. My child, if born and raised in Japan, will speak Japanese). We see, then, that a proximate process resulting in what a particular language spoken by a given population looks like does not necessarily speak to the evolutionary function. In other words, it is the device that allows the performance of Language that is adaptive, not the individual language itself.
One question being asked in the study of cultural transmission is why a particular language looks like it does, while we also know that there are 6000 different versions that perform the same (ultimate) function. I would even argue that asking how proximate processes shape languages is actually the most exciting and interesting avenue of inquiry precisely because it’s so blindingly obvious what the adaptive function of language is. But perhaps the value in this endeavour is somewhat neglected, in part, because of the same impression that Francis (1990) had: “the attitude, implicit in the term ultimate cause, [is] that these functional analyses are somehow superordinate to those involving proximate causes” which would be a shame. It seems to me that the coarse grain of ultimate vs proximate perhaps doesn’t do enough to help complex proximate study to position itself in the wider theoretical framework, and the best way to proceed from this might be to couch explanation in terms of Function, Phylogeny, Ontogeny and Mechanism. I think more fine-grained terminology grants us more explanatory power, in this case.
A final question in this debate that came up too many times during discussions with the LEC is: what does keeping the traditional paradigm “buy us”? Well, the first answer to this is consilience with one of the most successful and robust theories in science. The same sentiment has been communicated by Pinker and Bloom (1990), who said: “If current theory of language is truly incompatible with the neo-Darwinian theory of evolution, one could hardly blame someone for concluding that it is not the theory of evolution that must be questioned, but the theory of language”. Part of the reason this debate may have arisen is that studies of cultural evolution have used evolutionary theory as an incredibly fruitful way of analysing cultural processes, but additional acknowledgement about how cultural adaptation is different to biological adaptation may be necessary. This difference is an aspect of Laland’s paper (shown in Fig 2) that I think is important, as it’s part of the reason that more nuanced frameworks for cultural evolution are now needed. Without this widespread acknowledgement, cultural evolution may be considered an extension of biological evolutionary theory instead of a successfully applied metaphor. It seems to me that the side of this debate one falls on is well predicted by whether one subscribes to the former interpretation of cultural evolution or the latter.
Knowing which level of explanation current work pertains to is a valuable part of evolutionary exploration, and abandoning this in favour of an approach where proximate processes are explanatory ends to themselves may mean the exploration of Function and Phylogeny may suffer. That said, it is telling, I think, that even in seeking to abandon the proximate/ultimate distinction, we must still exploit this existing terminology in order to explain such a position. That natural selection has explained countless adaptations in all living things is certainly not trivial, and to reject the theory giving rise to ultimate explanations as they’re currently defined is to reject this fundamental aspect of evolutionary theory. The big problem seems to be that we’re coming to understand proximate processes as so elaborate and complex, that a more nuanced framework is needed to deal with the dynamics of those processes. I reckon, however, that such a framework can be developed within the traditional paradigms of evolutionary theory.
References
Francis, R.C. (1990) – “Causes, Proximate and Ultimate” Biology and Philosophy 5(4) 401-415.
Laland, K., Sterelny, K., Odling-Smee, J., Hoppitt, W. & Uller, T. (2011) – “Cause and Effect in Biology Revisited: Is Mayr’s Proximate-Ultimate Distinction Still Useful?” Science 334, 1512-1516.
Mayr, E. (1993) – “Proximate and Ultimate Causations” Biology and Philosophy 8: 93-94.
Millikan, R. (1993) – White Queen Psychology and Other Essays for Alice, Cambridge, Mass: MIT Press.
Pinker, S. & Bloom, P. (1990) – “Natural language and natural selection” Behaviour and Brain Sciences 13, 707-784.
Scott-Phillips, T. Dickins, T. & West, S. (2011) – “Evolutionary Theory and the Ultimate-Proximate Distinction in the Human Behavioural Sciences” Perspectives on Psychological Science 6(1): 38-47.
Sperber, D. & Origgi, G. (2000) – “Evolution, communication and the proper function of language” In P. Carruthers and A. Chamberlain (Eds.) Evolution and the Human Mind: Language, Modularity and Social Cognition (pp.140-169) Cambridge: Cambridge University Press.
Tinbergen, N. (1963) “On Aims and Methods in Ethology,” Zeitschrift für Tierpsychologie, 20: 410–433.