Time to
saddle-up and once more ride my current hobby horse, or one of them at least.
In this case, the idea that natural language is the simplest aspect of human
activity that is fundamentally and irreducibly computational in nature.
Let’s back into
it.
* * * * *
Is arithmetic calculation computational in kind?
Well yes, of
course. If anything is computation, that sure is.
Well then, in my current view, arithmetic calculation
is language from which meaning has been completely removed, squeezed out as it
were, leaving us with syntax, morphology, and so forth.
Elaborate.
First, let’s remind ourselves that arithmetic
calculation, as performed by writing symbols on some surface, is a very
specialized form of language. Sure, we think of it as something different from
language…
All those years
of drill and practice in primary school?
Yes. We have it drilled into our heads that arithmetic
is one thing, over here, while language is something different, over there. But
it’s obvious, isn’t it, that arithmetic is built from language?
OK, I’ll accept
that.
So, arithmetic calculation has two kinds of symbols,
numerals and operators. Both are finite in number. Numerals can be concatenated
into strings of any length and in any order and combination.
OK. In the
standard Arabic notation there are ten numerals, zero (0) through (9).
That’s correct.
And we’ve got
five operators, +, -, * [times], ÷, and =. And, come to think of it, we
probably should have left and right parenthesis as well.
OK. What’s the
relationship between these two kinds of symbols?
Hmmmm….The
operators allow as to specify various relationships between strings of
numerals.
Starting with,
yes, starting with a basic set of equivalences of the form, NumStr Op NumStr = NumStr, where Op is one from +, -, *, and ÷ and NumStr is a string of one or, in the
case of these primitive equivalences, two numerals. [1]
Thus giving us those tables we memorized in grade
school. Right!
What do you
mean by semantics being removed?
Well, what are the potentially meaning-bearing
elements in this collection?
That would be
the numerals, no?
Yes. What do they mean?
Why, they don’t
meaning anything…
Well… But they aren’t completely empty, are they?
No.
Elaborate. What’s not empty
about, say, 5?
5 could
designate…
By “designate” you mean “mean”?
Yes. 5 could
designate any collection with five members. 5 apples, 5 oranges, 5 mountains, 5
stars…
What about an apple, an orange, a mountain, a star, and
a dragon?
Yes, as long as
there’s five of them.
Ah, I see. The
numerals, or strings of numerals, are connected to the world though the
operation of counting. When we use them to count, they, in effect, become
numbers. But, yes, that’s a very general kind of relationship. Not much
semantics or meaning there.
Right. And that’s what I mean by empty of semantics.
All we’ve got left is syntax, more or less.
Sounds a bit
like Searle in his Chinese Room.
Yes, it does, doesn’t it?
The idea is that the mental machinery we use to do
arithmetic calculation, that’s natural computation, computation performed by a
brain, from which semantics has been removed. That machinery is there in
ordinary language, or even extraordinary language. Language couldn’t function
without it. That’s where language gets its combinatorial facility.
And THAT sounds
like Chomsky, no?
Yes.
* * * * *
And so it goes,
on and on.
When the
intellectual history of the second half of the twentieth century gets written,
the discovery of the irreducibly computational nature of natural language will
surely be listed as one of the highlights. Just who will get the honor, that’s
not clear, though Chomsky is an obvious candidate. He certainly played a major
role. But he didn’t figure out how an actual physical system could do it (the
question was of little or no interest to him), and surely that’s part of the
problem. If so, however, then we still haven’t gotten it figured out, have we?
* * * * *
[1] Isn’t that
a bit sophisticated for the Glaucon figure in this dialog? Yes, but this is a 21st century Glaucon. He’s got a few tricks up his
sleeve.
There is a “Skeptics In The Pub” event in Glasgow on March 4th, where Dr Thom Scott-Phillips will be discussing the perceptions and misconceptions of evolutionary psychology, in light of the public backlash against it that seems to be increasing all the time. This kind of public engagement is very sorely needed if we are to combat the rampant misinformation that crops up in both academic and non-academic communities. Among the criticisms being addressed at the event are the claims that evolutionary psychology is sexist, racist, or otherwise politically problematic. This is an important discussion.
From what I see around the feminist blogosphere, evolutionary psychology has a bad rap. Some recent examples I’ve come across include comments such as: “This new junk science named “evolutionary psychology” is the last variant of the male supremacy bible, following Freud’s mythology” and “[the way this article approaches the problem] is a bad idea [because] It smacks of evo psyche”. Even more liberal feminist blogs such as The F Word UK toe a similar line: Josephine Tsui seems to be on a personal mission against Evolutionary Psychology, armed with such ludicrous arguments as “You cannot replicate Evolutionary Psychology therefore it fails the methodologies of science” which display both an immature line of thinking and a fundamental misunderstanding of the theoretical motivations and methodologies entailed. Needless to say I’ve never seen this criticism leveraged against Evolutionary Biology, despite it being applicable to both.
Evolutionary psychology has a sound theoretical basis; it has been well established that natural selection is a means by which complex life and complex behaviour occurs. This tends to worry political movements like feminism, which has its roots in social constructionism. Such worry is unfounded; there is certainly a role for social constructionism within an evolutionary account of human behaviour. Put broadly, our plastic brains depend on complex social learning and pedagogy, which is an established cornerstone of human success. This ability to respond to (and be shaped by) the cultural environment has itself been selected for in humans, and can account for all manner of behaviours from language to mating preferences. Keep reading for a demonstration of how evolutionary psychology can in fact lend itself very well to the goal of engineering of social change.
So, on one side of the sexually selective understanding coin is a worried feminist movement, who risk losing a good grasp of evolutionary psychology by dismissing it entirely. On the other side, are the misogynist (mis)interpretations that have inspired this trepidation in the first place. That evolutionary psychology is abused and misinterpreted by misogynists and racists (and let’s be real here, this has happened a lot) is the problem, and it’s a serious one with real political consequences. Just this year, Steve Moxon submitted evidence to parliament (and was subsequently invited to speak) against the development of measures to encourage women in the workplace. Evolutionary psychology formed the backbone of his case, and he is not alone. Only an informed public can approach these claims with adequate discernment, so it is important that we address how some claims are morally wrong and incorrect. But it is also as important to discuss why they do not represent anything inherent about evolutionary psychology as a discipline.
We can illustrate the first way that evolutionary psychology can be wrong by using the problem of eugenics. Eugenics is theoretically sound, in as much as we know that we can selectively breed to a criteria and expect a predictable result; we’ve been doing it with dogs for 10 thousand years. This is also morally wrong and should not be attempted in humans. Just because eugenics is morally reprehensible, however, doesn’t mean we say the principles of artificial (or natural) selection aren’t true. Nor should this be the case for evolutionary psychology as a field; that it has been misapplied/misinterpreted within our social context (or just says something that we don’t like) simply does not speak to how scientifically correct it is. Another way that the interpretation of these studies can be grossly wrong is the Naturalistic Fallacy; the idea that if something is natural, it is inherently good or should be normative. This is obviously untrue; my human body is adapted to long-distance running, but I reject outright the idea that this is something I ought to engage in.
While citing the naturalistic fallacy is a good answer to most any claim about innate human proclivities, I think it’s also necessary to refute specific claims on their own grounds where possible. The final way for evolutionary psychology to be wrong is simply that rationalisation isn’t science, and instances where it is being passed off as such can be exposed for what they are. To illustrate, we generally do not dismiss the entirety of modern medicine as false because of the historical mistreatment of pregnant women in childbirth by doctors. Here, we can see that those occurrences are indeed morally wrong. However, it’s also the case that those instances are bad medicine by medicine’s own standards. Similarly, instances of bad science in evolutionary psychology, where latent misogyny and racism rears its head, can be refuted on their own grounds. This can and should be done without blithely dismissing the entire field.
It is a disaster that large factions of social justice movements are on the verge of outright anti-intellectualism when it comes to evolutionary psychology. Preserving ignorance about the field with out-of-hand dismissal neglects the potential for this tool to contribute to worthwhile political goals. We don’t have to stop at simply refuting the harmful instances of bad evolutionary study; there is also a positive agenda to be highlighted here. In the spirit of this, I’d like to share a preview of some work I’ve been hobbying with Justin Quillinan, inspired by a recent paper called “Asia’s Missing Women: A Problem in Applied Evolutionary Psychology?”. The paper aims to explore sex-preferential parental investment, which is a prolific problem in parts of Asia, the Middle East and North Africa, where the population’s sex-ratios are heavily male skewed as a result. It is already well documented that women suffer like this, so what can an evolutionary analysis can bring to the table? The problem, as presented in the paper, is this:
Asia’s missing women are, in economic terms, an aggregate outcome of millions of parenting decisions. The individual drivers behind those decisions emerge from interactions between our evolved parenting preferences and social and economic circumstances.
How do we untangle this seemingly nebulous problem? How do we determine why many self-perceived individuals act in such a similar way that the net result is the literal eradication of the female class? The approach of the paper seems to be largely influenced by economics, which is fairly central to a lot of evolutionary work. Game theory can be paraphrased as something like “given that the rules of the game are (x, y, z), which strategy should I employ to reap the most advantageous outcome?”. The most successful strategy is the one that is most likely to survive in the population, and hence it is the one we most expect to find. If we treat the problem of parents favouring boys over girls as a solution to the problem of parents’ circumstances, the question then becomes “what are the parameters that make this survival strategy worth employing such that it is so common?”
By comparing the commonalities in cultures that have this problem, Brooks identifies some ecomonic and social factors that may reward preferential parental investment. This is important: it means that campaigners for change don’t have to simply say the reason that girls are selectively aborted or neglected starts and ends with “girls are undervalued in these cultures”. Despite how true that is, it is also true of many cultures who do not have skewed sex ratios, and doesn’t really point to any concrete way of tackling the problem. If we can identify the driving factors that make parents behave this way with an evolutionary analysis, it means we can target specific structures with a specific end goal in mind.
In an interesting and wide-ranging investigation, the paper compares skewed parental investment occurence in non-human animals with the social and historical particulars that have led to this behaviour in disparate human populations. In doing so, Brooks proposes that male sex-biased populations are the systematic result of a population’s patrilineage, patrilocal kinship systems, and the dowry system. It was my hunch that the sex-biased population ratio could be reducible to patrilocality alone; that is, the system whereby women leave their blood relatives in order to live with (and care for) their husband’s family when they marry. Let us assume that the number of blood relatives in your family is a proxy measure of fitness. In a patrilocal social order, it is necessarily the case that having a son is more advantageous than having a daughter – precisely because daughters will always leave. Let us now also factor in the effects of infanticide and abortion; the option to neglect/kill your male/female offspring according to whether or not the most successful families you know had a boy or girl, will lead to the preferential elimination of females.
So we’ve implemented a model (source code available here) demonstrating exactly this:
We start with a population of agents separated into a number of families of a single breeding pair each single individuals. At each time-step, the following events occur: 1. Reproduction: fertile breeding pairs of agents have a new agent ‘child’ of random sex. A fertile pair is one that does not have an unmarried child that is younger than the age of maturity. 1.2. Abortion: Now, the pair can choose to either keep or to abort their new agent. To make this decision, they choose a random family, with larger families having a proportionally higher probability of being chosen. If the sex ratio of that family is a mismatch to the child they have just had, they will abort the child. Otherwise, they will keep it. (ETA: the abortion decision is based on the sex ratio of all the offspring of the chosen family). 2. Marriage: Every single, mature agent attempts to pair with a random, opposite sex partner from a different family, to form a breeding pair. In patrilocality, the female leaves her family and is appended to her husband-agent’s family to form a breeding pair. (In the matrilocality sanity check, the situation is vice versa and male agents join their wives’ families). 3. Death: Agents above a certain age are removed from the population. If a family no longer has any members, we generate a new breeding pair individual so that the population doesn’t die out.
The first null model was as above, with the omission of step 1.2. We later implemented one that is as above, but minus patrilocal marriage (ie, married agents simply form a new family pair) because this is a better comparison.
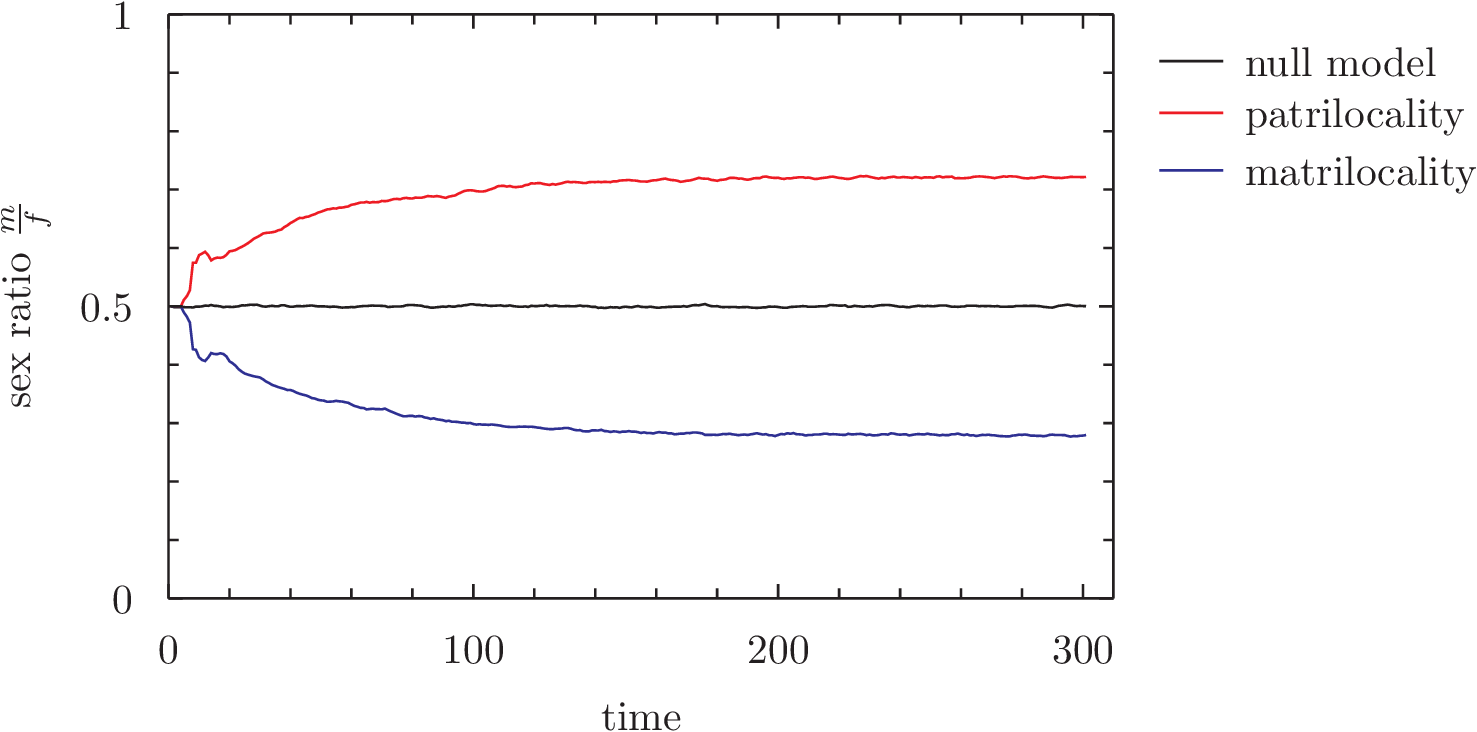
At each time-step, we measure the sex ratio of the population. This is what happens (wordpress is terrible, click for a clearer image):
Average of 500 runs, seeded from 50 family breeding pairs
The sex-ratio of the population is skewed in the direction of the sex that determines family locality – that is to say, patrilocality alone systematically results in the preferential abortion of female offspring, and a higher ratio of males in the population to females. This model will hopefully lend itself to some further work exploring the role of the dowry in maintaining the system by offsetting the costs of giving away offspring, preferential marrying, and how a shift toward “nuclear” family arrangements may have lifted the cost-benefit situations disadvantaging females (and thus making dowry systems redundant).
UPDATE (09/02/12): Here is the data using an amended null model:
Average of 1000 runs, seeded from 50 individuals
The ‘bump’ at around time-step 20 in the first graph noticed by Sean (see comments section) doesn’t appear here; this was an artefact of seeding with identical pairs that breed and die at the same time. Seeding with individuals has smoothed out that curve; staggering the ages of agents would likely smooth it out further. The extra noise in this model means that the skew is less pronounced than before; note the Y axis is zoomed in to ~0.3 – 0.7. The null model here is of the null hypothesis; abortion still happens, the only difference is that instead of a married agent appending to their spouse’s family, the married couple form a new family pair (ie. no matri/patrilocality). This means that any single given population’s sex ratio is susceptible to drift; early, small aberrations toward male or female will become magnified over time. This is, however, equally likely to happen for either male or female, and so the average of 1000 populations shown here is stable at 0.5.
Further implications:
An important additional observation in Brooks’ paper is an examination of the wider social consequences of this particular set of circumstances. The paper names elevated levels of “men competing furiously for wealth and status” as well as “risk-taking, violence, gambling, alcohol and drug abuse, kidnapping and trafficking of women, and the use and abuse of prostitutes” as consequences of surplus males in the population. The implication is that, by this model, these large societal problems can be addressed at least in part by balancing parental investment in children of both sexes, which would be remarkable.
At first blush, the idea that violence results from a surplus of men who don’t have a good enough chance at mating with women has some worrying and problematic implications. It is, nevertheless, intuitively true within a culture of male entitlement, which is something that feminists have long observed – that male violence is the result (and the maintenance) of a patriarchal social order. Since patrilineage, and patrilocality in family structure specifically, are identified as the preconditions for preferential parental investment in males, the eradication of this social order is a necessary step in redressing the sex-ratio balance. The end of patrilineal traditions and patrilocality are also a step toward dismantling a culture of male entitlement more broadly. As a direct consequence, then, this strategy dismantles the structures supporting male entitlement itself at the same time as addressing the skewed sex ratio, and does not simply consist of “giving the men more women to stop them fighting”.
It seems to me that a feminist account that names a culture of male entitlement as the cause for violent female oppression, and an evolutionary account that names structural entitlement systems as the cause for the mass devaluation/infanticide of female offspring are very much on the same page. This approach also very clearly illustrates the compatibility between evolutionary analysis and the socio-economic determinism that is fundamental to radical political thought, precisely by demonstrating how population-wide behaviour can directly result from external economic and social parameters, rather than some innately predisposed condition. We hope that this is at least one small demonstration of how evolutionary psychology and social justice can be rather natural allies.
A few months ago, a documentary I saw on the Discovery Channel covered some research by Graziano Fiorito and colleagues at the Stazione Zoologica in Naples. They were investigating observational learning in wild Octopus vulgaris with a puzzle-box experiment similar to those demonstrating cultural transmission in chimpanzees.
It goes like this: there’s a tasty and terrified crustacean running around in a perspex box that has two possible ways of being opened by hungry octopuses. The experimenters capture a wild octopus (let’s call him Steve) from the harbour (which I’ll get back to in a minute), and they put it in a tank with the puzzle-box. After Steve stares hopelessly at the box for a while, it is then removed from the tank. Steve the kidnapped octopus then gets to watch a captive octopus in the next tank being presented with the same puzzle-box containing the delicious crab. Of course, the captive octopus has been confronted with the puzzle-box enough times that it has worked out a successful solution, and so opens the box like a pro. Steve is then presented with a crab in a puzzle-box again, except this time he goes straight for the crab using the same solution he just learned from the captive octopus. Here is a clip from the documentary, showing a trial of this experiment (NB: not the best quality).
It turns out that the papers on this go back as far as Fiorito & Scotto 1992, and it seems this was the first time observational learning had been demonstrated in invertebrates. The reason I’m interested in reporting this is because the documentary I watched explained another possible motivation/interpretation for Fiorito’s work that I can’t find in any of his actual papers. The octopuses used in the experiments were all caught from the harbour at Naples just before the experiments, which of course controlled for any prior experience with the puzzle boxes. But the results were reported as particularly interesting because the Naples harbour had been overfished and disrupted, resulting in an increase in marine predators that eat the small octopus vulgaris as well as fish that the octopuses themselves rely on. These harsher environmental conditions resulted in the octopuses being forced to inhabit a smaller space alongside each other. As a result, young octopuses were frequently exposed to, and even coexisted with, older octopuses. This is a weird situation for an octopus; they usually live solitary lives and never even meet their own mothers, who die of starvation while caring for the eggs (the fathers die within a few months of having mated). The only real interactions are mating, and conflicts between rival males while competing for a mate.
Toward the end of the documentary, the voiceover growled against some dramatic music about how the combination of observational learning capacities and increased predation pushing octopuses into groups meant that it was only a matter of time before we’re overthrown by octopus vulgaris. This made me think of Dunbar’s “social brain hypothesis” for the emergence of language, and whether I really should prepare to welcome our new octopus overlords. Talking specifically about primates, Dunbar (1996) states that “[primates] in general exhibit two responses to increased predation: they grow physically bigger [or] they increase the size of their groups” (p.110). In order to maintain these groups, that are essential for survival in harsh ecological conditions, Dunbar suggests that standard primate grooming behaviour becomes too time consuming and costly in order to keep up with the rapidly expanding social group, creating a pressure for a more efficient method of bonding and communicating that allows the size of the group to continue increasing. Again with reference just to primate communication, Dunbar says “This [efficient mechanism] need not have involved any dramatic change, for as the studies by Seyfarth and Cheney have shown, primate vocalizations are already capable of conveying a great deal of social information and commentary.” (p.115) In addition to primates, and adding further credence to this idea, it’s been shown that the older Matriarchs of elephant groups make use of vocalisations to seemingly instruct their group on how to fend off lion attacks (McComb et al., 2011; previous Replicated Typo coverage here). It seems to me that the difference between the primates/elephants and the octopuses is that the former endeavour to actually enrich the environment from which their conspecifics extract information; they don’t just learn, they inform. As far as we can tell, there is no such communication like this – that is, enriching the environment in some way that helps other octopuses learn or survive – happening among the octopuses. That said, we know from mating displays and conflicts that cephalopods can communicate with chromatophore signalling. As an interesting aside that is reflective of their cognitive abilities and capacity for suffering, octopuses are treated as honorary vertebrates by UK animal testing laws.
It’s interesting enough that marine biologists (or at least those reporting on marine biologists) seem to have the same idea as Dunbar about the necessary preconditions for successful societies of animals, but why not let’s get wildly speculative? If (..!) existing in groups is in fact adaptive for these octopuses in the face of increased predation, and the competition between them for resources isn’t too great a counter-factor, it seems the only ingredient missing from an octopocalypse is the emergence of some cooperative behaviour. Someone should keep an eye on that harbour.
References
Dunbar, R. (1996) Grooming, Gossip, and the Evolution of Language. Harvard University Press: Cambridge, Massachusetts
Fiorito, G. & Scotto, P. (1992) “Observational learning in Octopus vulgaris” Science 256, 545-546.
McComb, K., Shannon, G., Durant, S., Sayialel, K., Slotow, R., Poole, J. & Moss, C. (2011) “Leadership in elephants: the adaptive value of age” Proceedings of the Royal Society B, published online.
We all tell stories. Be it in the elegant prose of a finely-crafted novel or merely relaying the day’s events to someone else, this capacity for storytelling is something found across our various cultures and has its roots deep in our prehistory. Stories are powerful tools with which we understand the world and share social information. They can also be dangerous. In the art of weaving together a tale, details might be omitted, others highlighted and some manipulated. Simple stories, where we abstract away from our complex and messy lives, are especially prone to this narrative fallacy, where we take facts and force an explanation into them, which, in the words of Nassim Nicholas Taleb, is misleading when it “increases our impression of understanding”. A similar warning is made in this brilliant video by Professor Tyler Cowen (of Marginal Revolution):
N.B. The featured picture is of the painting The Boyhood of Raleighby John Everett Millais. The story behind its use here is simple: it appears on the storytelling page of wikipedia. It also began me on a quest of hyperlink jumps, but that’s another story… Yawn.
Outside the world of evolutionary linguistics I used to spend some of my time working in a charity shop. Of the many dull moments, much of which spent bickering with overzealous bargain hunters about the arbitrary nature of our pricing, there were a few gems of conversation. On one of these days, I found myself conversing with several people about language change, when one of the customers chimed in with something I hadn’t heard before. He said, quite confidently, that the origin of speech and language lay in our ancestor’s proclivity for getting stoned. I humoured him on the magic mushroom hypothesis of speech origins, until he decided to share his wisdom about the foretold destruction of our society in 2012 (at which point I directed him to our copy of Emerich’s latest disasterpiece). Still, it appears he wasn’t completely barmy, at least on the speech origins front, as there is a Stoned Ape theory of human evolutionby one Terence McKenna (from Wikipedia):
The mushroom, according to McKenna, had also given humans their first truly religious experiences (which, as he believed, were the basis for the foundation of all subsequent religions to date). Another factor that McKenna talked about was the mushroom’s potency to promote linguistic thinking. This would have promoted vocalisation, which in turn would have acted in cleansing the brain (based on a scientific theory that vibrations from speaking cause the precipitation of impurities from the brain to the cerebrospinal fluid), which would further mutate the brain. All these factors according to McKenna were the most important factors that promoted evolution towards the Homo sapiens species. After this transformation took place, the species would have begun moving out of Africa to populate the rest of the planet. Later on, this theory by McKenna was given the name “The ‘Stoned Ape’ Theory of Human Evolution”.
I’m fairly sure this just offloads part of the craziness onto McKenna, but I might name drop the theory next time I’m looking for a more lively reaction when discussing language origins.
N.B. This is one of my many posts that was written some time ago. I decided to publish in 2012 just in case the customer was right about our impending doom. With that out of the way, we can now focus on the critical issues surrounding the size of a language’s phoneme inventory and the distribution of Psilocybe cubensis.